



从科学和逻辑的角度解释为什么武汉冠状病毒是人造的
文章来源:nerdhaspower.weebly.com于 2020-04-15
自从疫情发生以来,关于武汉冠状病毒的源头有非常多的争议。可以这么说,它的两种可能来源(自然进化或人造)似乎都有一定依据。对此全世界理应充分的分析和讨论。在社交媒体上,两种说法基本势均力敌。可是在科研论文或其他文章中,支持病毒来源于自然并谴责所谓“阴谋论”的占绝大多数。而支持另一边的,就是认为病毒来源于人造的,其科学论文或其他文章则几乎没有。在这篇文章里,我将用科学证据和逻辑推理来证明,这个武汉冠状病毒绝不是来源于自然。
我的分析会建立在确实可靠的证据之上。同时,我会主动排除不可信的,甚至是可疑的所谓证据。因为有些“证据”很有可能是中共主动掺进来,已达到混淆视听和掩盖真相之目的。鉴于此,我在分析中将扮演一个集科学家,侦探,和法官三重身份为一体的一个角色。
人们为什么觉得武汉冠状病毒的来源可疑
这种想法很大程度上源于对病毒序列的比较和分析。
一般有两种比较序列的方式,一个是比较基因序列,一个是比较蛋白质序列。对于病毒来说,这两种比较基本等同。这是因为病毒的基因序列基本全部被翻译成蛋白(病毒一般是一次性把自己的整个基因组翻译成一个长肽链,然后再用蛋白酶把它切成各个单独的蛋白)。所以我们在这儿就只用蛋白序列来比较几个相关的病毒。
通过这样的序列比较,很明显可以看出武汉冠状病毒和2003年的SARS冠状病毒有86%左右的一致性。这个程度的一致性基本就是说,两个病毒是同一类,但从进化角度来说,武汉冠状病毒肯定不是从SARS进化来的。这一点也是病毒领域里大家公认的。
值得注意的是,武汉冠状病毒与两个蝙蝠冠状病毒有极其诡异的相似性。这两个蝙蝠病毒(ZC45和ZXC21)统称为舟山蝙蝠病毒,因为都是从浙江舟山采集到的。从全序列的比较来看,舟山蝙蝠病毒和武汉冠状病毒一致性达到了95%。而对于序列里的绝大多数蛋白来说,这个一致性是普遍的,有的甚至更高,比如E蛋白的一致性是100%。Nucleocapsid蛋白是94%,membrane蛋白(膜蛋白)是98.6%,S2蛋白(spike蛋白的后半部分)是95%。然而,诡异的是,S1蛋白,也就是spike蛋白的前半部分,非常与众不同。在这里,两个病毒序列的一致性突然降到了69%。这种一致性的分布(所有其他部位95%,而仅一个特定蛋白69%)从遗传进化的角度来讲是极其诡异的。
说它诡异是因为自然进化绝大多数情况下是通过随机突变的方式进行的。那么假如仅仅是随机突变的话,祖先和后代之间序列上的差异是应该基本均匀分布的。或者说各个蛋白之间的一致性应该都差不多,不应该出现一个蛋白如此与众不同的局面。
那么有没有其他的进化方式会导致这样奇特的蛋白一致性的分布呢?是的,有一种方式可以做到,就是基因重组。我会在之后的段落里详细分析为什么天然的基因重组也不可能使舟山蝙蝠病毒或其近亲进化成为武汉冠状病毒。
现在,让我们把目光聚焦在这个与众不同的S1蛋白身上。
图1. 冠状病毒的模拟图。红色的就是spike蛋白。图片来源于美国疾病控制中心(CDC)
Spike蛋白就是装饰在病毒表面的红色的凸起(图1)。冠状病毒的名字就是从它而来,因为它使病毒在电镜下看起来就像皇冠。从功能角度来说,spike蛋白非同小可,它是冠状病毒的“钥匙”,可以打开我们(宿主)细胞的“锁”,从而进入并侵染细胞。
图2显示的是SARS的spike蛋白的结构。你可以把这样的结构想象为给真人拍的照片,它就是这个蛋白的真实面目。因为武汉冠状病毒和SARS冠状病毒的序列一致性也相当高,所以武汉冠状病毒的spike蛋白在结构上应该和这个SARS的spike基本一致。最近刚刚发表的文章也印证了这一点(1)。
图2. SARS病毒spike蛋白的结构以及它如何结合人体细胞上的受体ACE2。
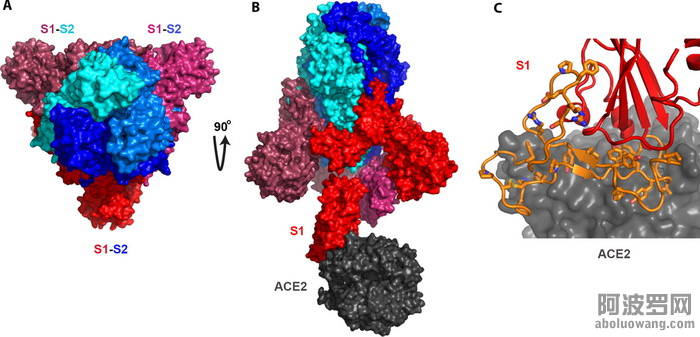
此图的制作是基于已发表的结构PDBID: 6acj(2)。A)三个spike蛋白形成三聚体,每个spike蛋白都由两个基本等长的部分组成,S1和S2。B)S2部分(蓝色)负责三聚体的形成,而S1部分决定和细胞受体的结合。在SARS这里,受体是人体细胞的ACE2。C)此图进一步揭示S1和ACE2结合的具体细节。橙色部分就是S1中和ACE2结合最关键的一段肽链。此橙色部分囊括了所有和ACE2结合的细节。其中最关键的氨基酸以棍状结构做了标识。此段橙色序列就是后文中提到的,如同被从SARS的spike蛋白中“复制”出来,然后“插入”舟山蝙蝠病毒的spike蛋白中去的关键肽段。这一“操作”直接可以导致一个新的,能感染人的病毒的产生。
如图2所示,三个spike蛋白聚集到一起形成所谓的三聚体。这个三聚体的形成完全取决于S2,也就是结构中的泛蓝色部分。这个S2可以认为是一个“钥匙”中用手抓着的那个部分。它不进入“锁”里,但对“钥匙”来说又不可或缺(此“钥匙”必须形成三聚体)。
Spike蛋白的另半部分就是S1,即图中泛红色部分。S1就是和细胞的受体结合的最关键的部分。你可以把S1想象成一把“钥匙”中真正进入“锁”的那部分。它必须和“锁”(细胞受体)的内部的精细结构严丝合缝才能把细胞的“门”打开。一个“锁”能不能被一个特定的spike“钥匙”打开完完全全取决于这个S1。换句话说,S1决定了一个冠状病毒会感染哪个宿主,哪种细胞。
了解了这些之后,你应该能更深刻的体会到为什么我之前用“非常奇怪”或“极其诡异”来形容武汉冠状病毒和舟山蝙蝠病毒之间序列对照的结果。当所有其他的部分都几乎一样的情况下,唯独这个决定了病毒侵染哪个宿主的S1发生了极其显著的改变。
好,我们再放大看一下具体是S1中的哪一小部分决定了和宿主细胞受体的结合(图2C)。在这里,和SARS的spike结合的受体是人体的ACE2蛋白。S1中对此结合起关键作用的这段肽链被标记为橙色,其中对结合ACE2起至关重要作用的氨基酸被标识成棍状结构。这段橙色肽链囊括了所有和ACE2结合所需要的微观结构。从下面的分析中你会发现,这段本是SARS的spike蛋白所独有的,并能充分决定spike蛋白和人体ACE2蛋白结合的肽段基本上被“复制”进了武汉冠状病毒之中。
图3. 相关冠状病毒的spike蛋白的序列比对。
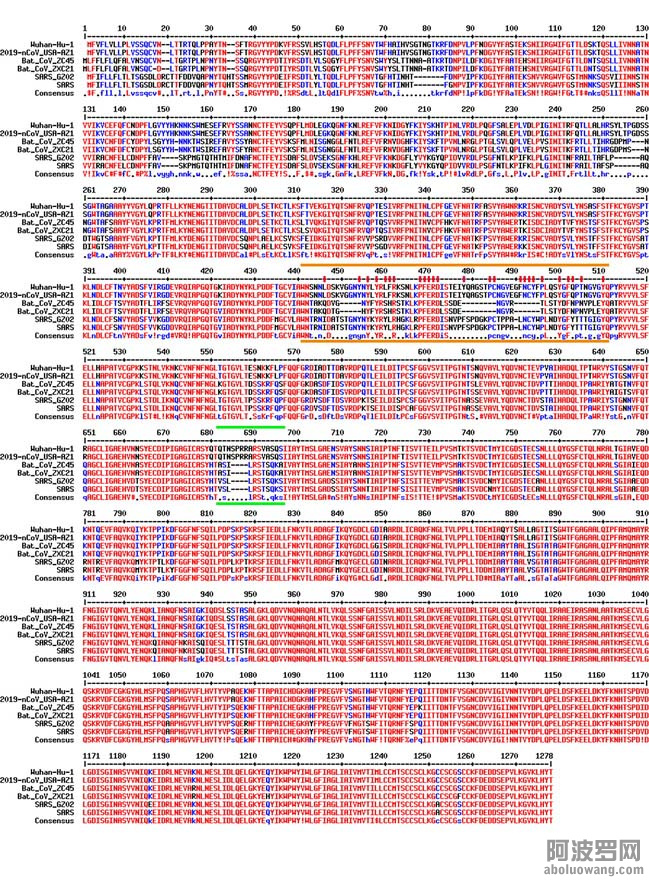
参与比对的包括两个是武汉冠状病毒(Wuhan-Hu-1, 2019-nCoV_USA-AZ1),舟山蝙蝠病毒(Bat_CoV_ZC45,Bat_CoV_ZXC21), 和SARS冠状病毒(SARS_GZ02,SARS)。两条橙色线之间的是和ACE2结合的关键区间。其中对结合起至关重要作用的氨基酸以红色竖杠标记。两条绿色线之间的是furin酶切位点。这样的位点只存在于武汉冠状病毒,而在其他任何的beta类冠状病毒中从来没有观察到过。序列对比用的是MultAlin webserver (http://multalin.toulouse.inra.fr/multalin/)。
图3是六个不同冠状病毒的spike蛋白的序列比对。其中,两个是武汉冠状病毒(Wuhan-Hu-1, 2019-nCoV_USA-AZ1);两个是舟山蝙蝠病毒(Bat_CoV_ZC45,Bat_CoV_ZXC21);两个是SARS冠状病毒(SARS_GZ02,SARS)。快速扫一眼,你就能发现对于spike蛋白的后半部分(690之后),也就是S2部分,六个病毒的序列都很接近。区别主要在前半部分(1-~690),也就是S1。假如你观察位于最上面的四个病毒的话,即两个武汉冠状病毒和两个舟山蝙蝠病毒,你会发现它们的S1也大体一致,唯有两三处明显不同。然而,就是这几处不同,尤其是不同里面的细节,基本上让我和很多其他人确信,这个武汉冠状病毒是人造的,而绝非来源于大自然。
第一个重要的不同之处就是图中用橙色线标注的地方。很明显,武汉冠状病毒在这一个区间内与舟山蝙蝠病毒非常不一样。与之形成反衬的是,在整个基因组的其他任何区域两种病毒都是高度一致的(超过 95% 的一致性)。与此同时,武汉冠状病毒的这一段却与SARS的spike蛋白的相关片段高度相似。注意,这个用橙色线标注出来的部分就是图2C中显示为橙色的肽段。像我们之前提到的那样,这部分肽段包括了所有和ACE2结合所需要的微观结构。一个很直观的感觉就是,SARS中这个最关键的片段被“复制”了,然后“粘贴”进了舟山蝙蝠病毒中,从而制造出了武汉冠状病毒。
再稍微仔细一点儿观察后,你可能会发现武汉冠状病毒和SARS冠状病毒在这一段似乎还有相异之处,因此又不是很明显的“复制”和“粘贴”。可是,当你非常仔细地在微观结构层面分析SARS的spike蛋白如何与人体ACE2蛋白结合之后(图2C),你会发现S1中所有对结合ACE2至关重要的氨基酸(图2C中以橙色棍状标注的,同时也是图3中用红色竖杠标注的氨基酸)都基本被精确保留了或者是替换成了性质类似的氨基酸。另一方面,两种病毒的spike蛋白在这一区间的不同之处基本全在不影响与ACE2结合的位置上。仅从序列上的这种信息就可以判断,武汉冠状病毒不仅能与ACE2结合,而且其结合的方式与SARS的spike蛋白结合ACE2的方式决无二致(图2BC)。
而舟山蝙蝠病毒,因为它们的spike蛋白缺少很多关键的氨基酸,它们与ACE2结合进而侵染人体的可能性是非常非常小的。
我们再梳理一遍武汉冠状病毒和舟山蝙蝠病毒的序列比较:两者在绝大部分蛋白上都几乎一模一样(95%一致性),而唯独在spike蛋白的S1这里,一段SARS的spike蛋白中专门决定与人体ACE2结合的序列被生生的“继承”了过来,替换掉了的是原舟山蝙蝠病毒中一段无法与人体ACE2结合的肽段。而且这个还并不是生搬硬套的继承:所有的关键氨基酸被保留而不参与结合ACE2的很多氨基酸都变了。这种方式的“替换”不可谓不神奇,而这个神奇“替换”的结果就是武汉冠状病毒“获得”了感染人的能力。此能力是舟山蝙蝠病毒所不具备的。
自然进化能如此精准,同时具有如此的隐蔽性吗?
假如以上还不算触目惊心的话,我们再看看这里面的第二个“魔术”。这次关注的焦点是图3中两条绿线之间的部分。在这里,唯有武汉冠状病毒有一小段额外的序列,就是SPRRA。有了这个独特的序列后,武汉冠状病毒的spike蛋白就能够在这个位置被人体的furin蛋白酶剪切。而此种剪切是被证明可以增强流感病毒(含有类似的spike蛋白)的感染力的。需要注意的一点是,除了武汉冠状病毒病毒之外,自然界中还没有发现任何处于同一谱系(lineage B)中别的beta类冠状病毒有这样的furin酶切位点。
进一步解释为什么这些序列上的变化不可能来自自然进化
我们之前简短地解释了为什么随机突变不可能导致一个类似舟山蝙蝠病毒的祖先进化成为武汉冠状病毒。现在我们再进一步地说明这一点。确实,相对于其他部位来说,冠状病毒的spike蛋白更容易出现变异。然而,当两个病毒的S1差异很大时,必然是因为它们在很早很早之前就分开进化了,并且进化过程中频繁地随机变异以适应不同的宿主(因为S1是决定侵染什么宿主的)。这种情况下,不光是两种病毒的S1部分一致性低,它们整体序列一致性一定也是偏低的。而现在的情况是武汉冠状病毒和舟山蝙蝠病毒全序列一致性高达95%,显示这两个病毒是非常近的近亲。在这个前提下,出现S1蛋白一致性只有69%的现象是完全不可思议的。所以说,武汉冠状病毒的S1蛋白绝对不可能是由舟山蝙蝠病毒的S1蛋白通过随机变异的方式进化而来的。
好,那我们再来说一下为什么武汉冠状病毒也不可能以基因重组的方式进化而来。
基因重组是指一个长基因中的某段可以被其他基因中与之类似的一段置换。基因重组也是自然进化的方式之一。它发生的频率远远低于随机突变。可它一旦发生,往往会给基因组里的某些位置带来非常大的,有时甚至很神奇的变化。
假如是自然进化中的基因重组导致了武汉冠状病毒这个神奇的存在,它会怎么发生呢?首先,这样的基因重组要想发生必然是一个类似舟山蝙蝠病毒的祖先和另一个冠状病毒同时侵染了一个动物体的同一个细胞。在这个前提下,非常偶然的会出现我们需要的基因重组,就是这个祖先病毒的某一段被另一个冠状病毒中的相似片段所替换。非常重要的一点是,从一个舟山蝙蝠病毒那样的祖先出发,最终进化出武汉冠状病毒的话,一次基因重组是不够的,必须有两次基因重组才可能进化出如此的结果。其中一次,这个祖先病毒必须和一个类似SARS的冠状病毒共存,然后通过基因重组精确地得到S1蛋白中专门和人体ACE2结合的那一小段(图2和图3中以橙色标记的部分)。除此之外,在另一个契机中,这个经过以上“改良”的祖先病毒还要再和一个在S1和S2之间包含furin酶切位点的冠状病毒共存于一个细胞,并通过基因重组将这个furin酶切位点准确地置换了进来。
另外,像我们提过的那样,基于舟山蝙蝠病毒和武汉冠状病毒高度的一致性(95%),我们可以合理推测出两种病毒并不是远古之前分开进化的。也就是说,以上提到必须发生的两个基因重组也都应当是相当近的年代发生的。
好,我们知道SARS的出现并感染人其实是进化中非常偶然的一件事。这也说明,要想自然中再出现一个和SARS类似,能侵染人,既结合人体ACE2,同时S1里的关键序列又和SARS不太一样的冠状病毒应该是更偶然的一件事。再让这样的一个类SARS病毒与舟山蝙蝠病毒共存并发生精确的基因重组,那几乎就是不可能的。除此以外,furin酶切位点在已知的同一谱系(lineageB)的beta类冠状病毒的spike蛋白中都是不存在的。虽然类似的furin酶切位点在相隔较远的冠状病毒中被发现过,但没有一个位点的氨基酸序列和武汉冠状病毒中的这个一模一样。也就是说,让舟山蝙蝠病毒和另外一个含有如此序列的furin酶切位点的冠状病毒共存于一个细胞,并通过基因重组将此furin酶切位点置换进自己的基因组,其可能性也是非常非常小的。
那么好,让这两个几乎不可能的事都发生的可能性是多少?我的答案是没门儿,现实中没有可能。因此,武汉冠状病毒绝对不可能来自于自然。
为什么有些文献必须被排除在外
有些关注了最近发表的科学论文的人可能会说,以上的分析是不对的,因为某个非常重要的发现并没有被考虑进来。而恰巧,这个发现是与我的论点相悖,支持病毒来自自然进化的。那我为什么敢对它完全无视呢?
简单一点儿的答案就是,我认为这个发现, 或者说“证据”,很有可能是伪造的。
请允许我现在转换我的角色,从一个科学家变成一个警探或者法官。假如我们把这个事件当成一个正在被调查或庭审的案件的话,那我们这里有一个最大的嫌疑人,就是来自武汉病毒所和P4实验室的冠状病毒专家,石正丽博士。从武汉疫情爆发刚一开始,就不停的有人怀疑是石正丽人工制造了这个病毒,并有意或无意地把它释放了出去。石一直是最大的,甚至唯一的嫌疑人。有意思的是,石在几周前在自然杂志(nature)发表了一篇论文(3)。里面她将当时刚刚公开不久的武汉冠状病毒的序列与其它beta类冠状病毒的序列做了比较,并由此描绘了武汉冠状病毒的可能进化路径。文章里报道了一个全新的蝙蝠的冠状病毒,RaTG13。此RaTG13与武汉冠状病毒有非常之高的一致性。这里最神奇的是,这个高一致性(98.5%)或者说非常低的变异率是均匀遍布于基因组的各个部分或者说各个蛋白的。我们之前怀疑武汉冠状病毒来源于人造,其主要根据是武汉冠状病毒与舟山蝙蝠病毒之间序列的一致性分布非常诡异,无法用自然进化解释。这个问题在RaTG13身上则完全消失了。RaTG13的spike蛋白和武汉冠状病毒的spike蛋白也高度一致(97.4%),与两个病毒间在其他部位的“变异频率”是一样水平的。第一眼看过去,这个RaTG13很明显就是与武汉冠状病毒同源的,两者不久之前有一个共同的祖先,从那分支并各自进化而来。这一“发现”有力地证明武汉冠状病毒是来源于自然的。这篇报道了RaTG13的论文就是被我完全无视,在之前的分析中故意忽略的“证据”。
根据可靠的消息来源,石正丽跟几个人分别承认过,她手中并没有真正的RaTG13的毒株。据她说,她的实验室是在七年前从云南的蝙蝠的粪便中分析基因片段,从而寻找可能存在的冠状病毒时发现的这个RaTG13的基因。用稍微通俗点儿的话说,事实上她是没有实物证据能证明RaTG13存在的,她只有这个病毒的基因序列,也就是一个由ATCG四个字母以各种方式组合出来的长链。
这个序列可不可能被伪造呢?这个其实再容易不过了。让一个人坐那儿往一个word文档里打字就可以了。一共不到3万个字母,不到一天就完活儿了。要是你还有一个模版序列,它和你想要得到的序列有98%是一摸一样的,那这个工作就要更容易一千倍了。新序列打完之后,只要上传到网上公开的基因数据库就可以了,期间没有什么严格的审核。上传了的序列就可以被公开引用,并拿来分析数据和发表文章了。
那么,现在的问题就是,这样一个RaTG13的序列能不能拿来当作证据呢?别忘了,这个事很核心的一点就是到底是不是石正丽本人制造了这个病毒。假如真是人为制造的话,那么这可以说是人类有史以来最大的犯罪,而石是这里唯一最大的嫌疑人。这种情况下,石是不是有为自己掩盖罪行的动机呢?假如她在自证清白时所用的证据不过是她刚刚在word文档中打出来的一连串字母,法官,陪审团,受害者,或者任何人应不应该认可这样的证据呢?
非常不幸的是,最近发表的很多科研论文都把分析和推理建立在这个RaTG13的序列之上。它们的结论也自然毫无意外:武汉冠状病毒一定来源于自然进化。我希望读者们能认识到或认同我的看法,就是这个RaTG13的序列很可能是石正丽伪造的,所以以此为依据得到的任何所谓科学判断都需要打个大大的问号。
好,我们现在再换一个角度看。这个RaTG13病毒的序列是很震撼的。任何这个领域的专家只要瞥一眼它的spike蛋白的序列就会立马意识到这个病毒和SARS很像,很可能可以结合人体ACE2,所以它非常可能具有感染人的能力。石本人就是这样的专家。石正丽实验室对外宣称,她们研究的目的就是希望通过对蝙蝠的冠状病毒的充分了解达到能够预警人类的作用,防止类似SARS那样的健康灾难的发生。那么本着这样的心态,石正丽怎么会在七年之间似乎完全忽视了一个像RaTG13这样的病毒呢?她怎么能忍七年而不发表这个惊人的发现呢?为什么只是在武汉疫情爆发之后,当人们开始怀疑武汉病毒的来源的时候,石突然决定发表这个RaTG13的序列呢?
所有这些都不符合常理。这些事实放在一块儿只能让人更加地怀疑石正丽。她或者直接参与了制造这个病毒,或者在帮忙掩盖真相,或是两者兼而有之。另外,这些事实当然也更加说明,这个RaTG13的序列必须被排除于任何的科学分析之中。
同样需要排除在外的是所谓穿山甲是病毒从蝙蝠传播到人的中间宿主这一说法。二月初的时候,华南农业大学的三位研究人员召开了一个新闻发布会,声称他们最新的科研结果显示穿山甲很可能是病毒的中间宿主。首先,这个新闻发布会召开的时间点就很可疑,当时坊间开始讨论说蝙蝠病毒不可能直接传染人,需要有一个中间宿主(因为病毒的spike蛋白需要在一个有类似于人的ACE2的动物中“学习”和适应如何与这样的ACE2结合)。以往的经验告诉我们,当什么事情必须存在才能让中共自圆其说的时候,这个事情一定会神奇的出现或者被某个中共控制的人“亲口证实”,就像石正丽的RaTG13。这次从天而降的是穿山甲的冠状病毒(从而佐证穿山甲是中间宿主)。这些科研人员都等不到发文章了,直接表示他们有证据:一个序列(又是序列)。具体来说,是他们发现的穿山甲冠状病毒的S1蛋白的受体结合区域(也就是和ACE2结合的区域,英文简称RBD)的序列信息。和之前一样,没有活体病毒,只是序列信息,而且当时还没公布。像我们在讨论RaTG13时提到的,这样的序列信息在word文档里打字就可以了。这次这个RBD更短,几分钟就能打完了。基于这种情况,这个穿山甲冠状病毒的序列信息同样有很大的可能性是中共为混淆视听而伪造的。
幸运的是,领域里人似乎也很快就放过了穿山甲。最终这些穿山甲冠状病毒的序列信息被两个科研组(华南农大和香港的管轶的科研组)分别发表并公布出来(4,5),可是这些序列却没能让业界相信穿山甲是中间宿主。这其中有一部分原因是穿山甲冠状病毒的spike蛋白序列中也没观察到furin酶切位点。
所以,就像RaTG13一样,最近这些证实穿山甲为中间宿主的科研论文也应该在分析时被排除在外。事实上,就在刚刚,这几个华南农大的研究人员向媒体承认,在对穿山甲冠状病毒的全序列分析之后,他们认为自己之前的说法有误,穿山甲不是武汉冠状病毒可能的中间宿主。
一些应该被高度关注的科研文献
我们刚刚花大篇幅解释了为什么某些“科研证据”必须被排除在外。现在我们再来看看某些其他的文献和证据必须被高度关注和认真参考。
第一,这两个与武汉冠状病毒具有诡异相似性的舟山蝙蝠病毒(ZC45和ZXC21)是由中共的军事科研单位收集并发现的。他们把科研结果,其中包括舟山蝙蝠病毒的序列信息,在2018年发表了(6)。在这里我想强调两点:一,如果武汉冠状病毒是人造的,那它一定是用舟山蝙蝠病毒为模版(template)做的;二,这篇文章揭示全世界只有中共手中有舟山蝙蝠病毒。
第二,石正丽在2015年与人合作发表了一篇Nature Medicine的文章(7)。他的合作者是北卡大学的Ralph Baric教授。这篇文章揭示了如何把一个普通的蝙蝠冠状病毒变成一个能侵染人的病毒。具体来说,他们把一个原本对人无感染能力的蝙蝠冠状病毒的spike蛋白换成了能够与人体ACE2结合的一个spike蛋白。这一个改变就把原来的蝙蝠病毒变成了针对人的冠状病毒。好,现在我们回头看看,在武汉冠状病毒发生的事基本就是按着这篇文章的脚本儿来的。现实中,并不是整个的spike蛋白被替换了,而只是替换了spike蛋白的S1中最关键的一部分。这个最小程度的改变却充分地给了被改造后的病毒感染人的能力。这里真正的不同之处是,这种微妙地改变spike蛋白的方式让试图找寻病毒来源的人们更容易迷失方向。
第三,一篇2006的文献揭示,以人为的方式在SARS病毒spike的S1和S2之间插入一个furin酶切位点之后,病毒和细胞膜融合的能力出现了大幅度的提高(8)。虽然如此操作并没有像预期的那样让这个实验室病毒出现超强的感染力,但已经公认的是存在furin酶切位点的流感病毒的致病性比没有这样位点的流感病毒高出很多。神奇的是,这个操作也如出一辙地出现在了武汉冠状病毒的spike蛋白之中(图3中绿线标注的肽段)。另外,拥有furin酶切位点的流感病毒通常可以感染更多类型的细胞,因而攻击的器官往往不局限于肺。假如你关注了一些最近有关武汉冠状病毒的病症特征的话,你可能已经注意到了,几个独立的报道已经描述了这个病毒不但攻击肺,同时也能攻击肝,心,血管,甚至中枢神经系统等等。
清晰的证据链和逻辑链显示中共为制造武汉冠状病毒的元凶
假如你把以上这些信息稍作整理,你会发现假如是中共制造了武汉冠状病毒的话,这件事是多么的符合逻辑。很明显,制造这个病毒所用的的模版,也就是舟山蝙蝠病毒,只有中共手里才有(6)。在这个基础上,他们要做的就是改变这个蝙蝠病毒的某些地方,让它变得能感染人,而且最好是传染性极强。而实际上发生的就是他们完全按着两个发表过的概念做出了这个新病毒(7,8):1)他们依照SARS蛋白结合ACE2的方式对舟山蝙蝠病毒的spike蛋白做了精确的改造,使之能按照SARS的spike那样的方式结合人体ACE2。如此一来,新病毒变得能感染人了;2)他们在spike蛋白的S1和S2之间插入了一个可以让furin酶来切割的位点,而此操作有可能让病毒增加致病力以及传染力。这两个概念可以说是中外文献中能找到的,仅有的两个如何改进冠状病毒的方式。结果,两者在武汉冠状病毒的身上都被完美的体现了。假如是大自然制造了这个病毒,那大自然一定非常仔细地研究了近些年人类在这方面的科研成果,并且把人类的几个最重要的发现极为忠实地应用在了她的工作之中(2,6,7,8)。
我们也可以再往回想一想。为什么中共治下有那么多的科研单位这么努力地在各处寻找冠状病毒呢?事实真是像他们说的那样,是为了更好地了解冠状病毒各种可能性,进而更好地预测未来可能由新的冠状病毒导致的疫情吗?那他们为什么不把精力花在制造广谱的冠状病毒疫苗或者是针对冠状病毒中的某个保守性高的药物靶点而制药呢?后者不但对人类健康更有直接意义,而且比预测哪个冠状病毒未来会爆发要容易不知多少倍。
另外一种可能性,就是他们收集这些野生动物的冠状病毒是为了制造生物武器。到底哪一个是真相?你可以自己判断。
对我来说,答案再明显不过,这次的武汉冠状病毒就是中共所制造的生物武器。
基于以上罗列的事实和对这些事实的分析,一个最合理的结论就是,除非中共能提供有效的证据证明自己是清白的,否则全世界已经有足够的理由相信武汉冠状病毒就是中共人工制造出来的。
引文出处:
1. Daniel Wrapp NW, Kizzmekia S. Corbett, Jory A. Goldsmith, Ching-LinHsieh, Olubukola Abiona, Barney S. Graham, Jason S. McLellan. Cryo-EM Structureof the 2019-nCoV Spike in the Prefusion Conformation. Science. 2020.
2. Song W, Gui M, Wang X, Xiang Y. Cryo-EM structure of the SARScoronavirus spike glycoprotein in complex with its host cell receptor ACE2.PLoS Pathog. 2018;14(8):e1007236.
3. Zhou P, Yang XL, Wang XG, Hu B, Zhang L, Zhang W, et al. A pneumoniaoutbreak associated with a new coronavirus of probable bat origin. Nature.2020.
4. Kangpeng Xiao JZ, Yaoyu Feng, Niu Zhou, Xu Zhang, Jie-Jian Zou, Na Li,Yaqiong Guo, Xiaobing Li, Xuejuan Shen, Zhipeng Zhang, Fanfan Shu, Wanyi Huang,Yu Li, Ziding Zhang, Rui-Ai Chen, Ya-Jiang Wu, Shi-Ming Peng, Mian Huang,Wei-Jun Xie, Qin-Hui Cai, Fang-Hui Hou, Yahong Liu, Wu Chen, Lihua Xiao, YongyiShen. Isolation and Characterization of 2019-nCoV-like Coronavirus from MalayanPangolins. bioRxiv. 2020.
5. Tommy Tsan-Yuk Lam MH-HS, Hua-Chen Zhu, Yi-Gang Tong, Xue-Bing Ni,Yun-Shi Liao, Wei Wei, William Yiu-Man Cheung, Wen-Juan Li, Lian-Feng Li,Gabriel M Leung, Edward C. Holmes, Yan-Ling Hu, Yi Guan. Identification of2019-nCoV related coronaviruses in Malayan pangolins in southern China. bioRxiv.2020.
6. Hu D, Zhu C, Ai L, He T, Wang Y, Ye F, et al. Genomic characterizationand infectivity of a novel SARS-like coronavirus in Chinese bats. EmergMicrobes Infect. 2018;7(1):154.
7. Menachery VD, Yount BL, Jr., Debbink K, Agnihothram S, Gralinski LE,Plante JA, et al. A SARS-like cluster of circulating bat coronaviruses showspotential for human emergence. Nat Med. 2015;21(12):1508-13.
8. Follis KE, York J, Nunberg JH. Furin cleavage of the SARS coronavirusspike glycoprotein enhances cell-cell fusion but does not affect virion entry.Virology. 2006;350(2):358-69.
