随遇而安
人生境遇不可选择,心态可以选择



机理这两个字简单, 但是内容很广很深, 需要无数科学家长年的努力解决,甚至是几代人的辛勤工作的结果。 葡萄糖转运机理就是这样。 这个机理大致可以简化为两个问题:
一。 为什么“只”转运葡萄糖? (注:为叙述简单, 这里用词不够准确)
二。怎么转运?
这是两个相对独立的问题。 可以理解不同的元件组合可以实现不同的功能。 就像丰田汽车既可以装Michelin的轮子, 也可以装Goodyear的轮子。 本田也可以装这两种轮子。 不是说两者完全没有交集, 但是至少现在的研究还没有深入到这个程度。
一, 为什么“只”转运葡萄糖?
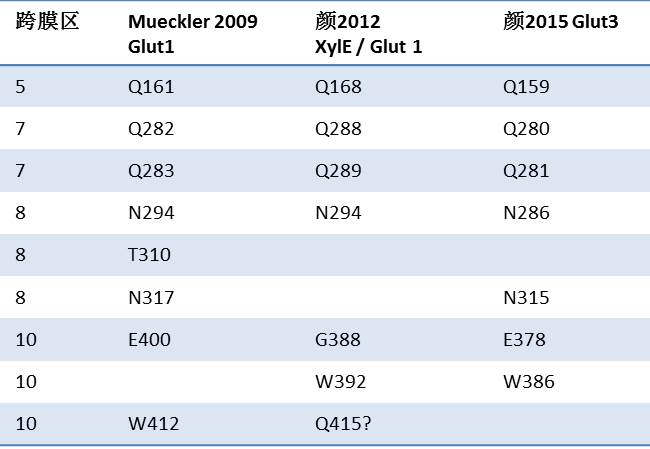
科学地讲,就是转运蛋白的底物特异性。 这个问题简单回答就是, 蛋白质需要在特定的位置有特定的氨基酸。 这些位点一般比较“保守”--即功能相似但不同的蛋白基本都有这些或者性质类似的氨基酸。 2009年Meuckler对GLUT1的研究就总结了这个糖蛋白的结合位点。 大部分在颜2012与2015都得到了证实(见下表), 但也有被晶体结构“否定”或 质疑的,这反映了大家某些地方还在“摸象“和科学进步中的不确定性。再下面的课题有两个。 1。为什么这些氨基酸决定了蛋白的特异性? 第二个问题是蛋白质与葡萄糖结合后如何发动转运。 现在给读者留一个小题目:为什么这个表里没有颜2014?
二 转运蛋白是怎么转运的
上述的底物结合位点并非转运蛋白特有,其他的蛋白特别是各种酶最先研究的就是底物的结合位点。 然而, 转运的机制是转运蛋白所特有, 也是阎颜之争的焦点。但不是葡萄糖转运蛋白所特有。 比如MFS超家族的蛋白转运的基本原理上是差不多的。
转运蛋白这个领域公认的基本模型就是1966的经典模型。 这个模型给转运机理的基本描述: 蛋白面向细胞外--〉蛋白抓住底物--〉蛋白面向细胞内--〉底物释放到细胞内--〉蛋白面向细胞外, 如此循环反复。 蛋白轮流地面向细胞外与细胞内,所以叫Alternating Access Model.
下面的工作有两个:
(1)。为这个基本模型提供证据, 于此同时,很可能排出其他模型。
(2)。提出更细节的工作原理,搞清楚转运蛋白是如何循环反复的?
这两项工作其实是交织在一起的。下面具体谈第二个问题:蛋白是如何从冲外变成冲里循环反复的?
如下图所示, Forrest 在2011年综述总结了两个。 一种是Rocker Switch. 就是蛋白质的蓝绿两扇门一开一合, 实现对外对内的轮流。 这一模型的要点是葡萄糖在蛋白里的结合位点在“纬度”上是不动的。
另一种是Gated Pore. 注意这个模型虽然与Channel有点象但是是一次一个的”thin channel”. Alternating Access通过外门与内门的轮流开关实现。 第三种机理是蛋白某一部分变形做 “搬运工”。 这一机制没有被Forrest总结。
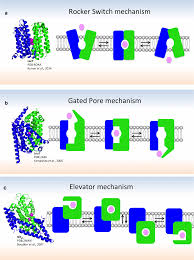
而Rocker Switch与 Gated Pore 并不是完全互斥的。 LeuT似乎就显示了两者的特质。 而Glut1-4 似乎也既有rocker switch 也有gating pore的机制。 “转运机理”这个词实际上是非常复杂的。 科学家用模型来简化现实的复杂性, 使得复杂的现实更容易被人理解, 但是不能说模型就能代表现实。 我想这也许是颜宁不提Rocker Switch与Gating Pore的原因吧,以免落了窠臼。 比如Glut 里的ICH这个门闩就在两个机制里不存在。
科学的进步是从粗浅到深入,从模糊到清晰慢慢地进步的。 1966 经典根据动力学的研究有了很粗糙的模型, 逐渐进步到用rocker switch 与gating pore等解释Alternating Access. 现在又多了门闩。 并且由于有了晶体结构清晰的摆拍,对机理的解释到达了一个一个氨基酸如何互相作用的细致程度,是生化实验无法做到的。 而这只是机理的一个侧面。 另一个侧面是symporter, unitporter, antiporter coupling的区别。 “葡萄糖转运机理的发现”是个万里长征的大工程,很难归功于一个人的功劳。
